Диагностика и ремонт датчика положения дроссельной заслонки / Блог им. Girman / OpenECU
В представленной статье будет рассмотрено устройство датчика положения дроссельной заслонки, диагностика и симптомы неисправностей ДПДЗ, а так же его ремонт.
Устройство датчика положения дроссельной заслонки
Итак, если Вы задались вопросом, каким образом устроен датчик положения дроссельной заслонки, то стоит сначала рассмотреть принцип его работы.
Датчик положения дроссельной заслонки относится к типу датчиков резистивного типа. Данное название обуславливает принцип его работы, а именно, если разобрать данный датчик, то внутри мы обнаружим подвижной элемент в виде ползунка, который скользит по дорожке в виде дуги или подковы. К одному из концов данной дорожки подается питающее напряжение, другой конец дорожки соединен с массой, а с подвижного ползунка снимается выходной сигнал.
Неисправность датчика положения дроссельной заслонки:
Какие же неисправности датчика положения дроссельной заслонки чаще всего встречаются на практике? Если отбросить неисправности связанные с перетертыми проводами, подходящими к датчику и т.
На датчик подается напряжение
В отдельных моделях автомобилях могут применяться датчики положения дроссельной заслонки с инверсной выходной характеристикой, то есть напряжение при закрытом дросселе будет максимальным, а по мере открытия дросселя оно будет падать.

Так же следует обратить внимание, что на автомобилях, где положение дроссельной заслонки задаётся при помощи электропривода ( в народе известная, как «электронная педаль») в указанных моделях положение дроссельной заслонки определяется при помощи не одного, а сразу двух потенциометров которые объединены в одном устройстве. При этом не имеет значения задает ли электронная педаль положение только в режиме холостого хода или во всем диапазоне. Один из двух потенциометров имеет инверсную выходную характеристику, а второй прямую выходную характеристику. На подобных системах, так же можно встретить концевой микро-выключатель который срабатывает в момент, когда педаль акселератора полностью отпущена водителем.
Как обнаружить неисправность датчика положения дроссельной заслонки без разборки датчика и снятия его с автомобиля:
— неисправность датчика положения дроссельной заслонки можно легко определить при помощи сканера, мотортестера или простого мультиметра.
 В данной статье мы рассмотрим пример обнаружения неисправности при помощи сканера.
В данной статье мы рассмотрим пример обнаружения неисправности при помощи сканера.Итак, зайдите в сканере в режим снятия параметров в режиме реального времени, после чего перейдите в раздел снимающий показания положения дроссельной заслонки в процентном соотношении или вольтаж на датчике, после этого начните
Не обращайте внимания на незначительные изменения осциллограммы, это может быть обусловлено дрожью Вашей руки. Так же следует отметить, что при низкой скорости обмена между сканером и ЭБУ автомобиля возможен пропуск дефектного слоя резистивной дорожки, если он совсем короткий, но данный факт скорее исключение, чем правило.

При снятии датчика с автомобиля так же не будет лишним осуществить промывку дроссельного узла, отложения на стенках которого, так же могут мешать нормальной работе датчика.
Ремонт датчика положения дроссельной заслонки
Восстановить изношенный резистивный слой на дорожках, в бытовых условиях невозможно, поэтому единственным способом ремонта без замены датчика или дорожек является возможность в некоторых датчиках смещения резистивных дорожек относительно ползунка. Для этого в датчике предусмотрен специальный винт который фиксирует то или иное положение дорожек относительно ползунка, поэтому допустим при сильном износе начала резистивного слоя дорожки мы можем, ослабив винт, сместить его в область недоступную ползунку и таким образом избежать замены датчика положения дроссельной заслонки.
Симптомы неисправности датчика положения дроссельной заслонки
В случае износа резистивного слоя, в зависимости от места износа автомобиль может вести себя различными способами.

Так же в отдельных случаях при замене оригинального датчика положения дроссельной заслонки на некачественный аналог может наблюдаться зависимость работы датчика от температуры, то есть по мере нагревания корпуса ДПДЗ выходное значение будет меняться. К примеру, на холодном двигателе датчик имеет выходное напряжение около 500 мВ, ЭБУ сохраняет данное значение, как положение закрытого дросселя и приступает к стабилизации оборотов холостого хода. После нагревания корпуса датчика, выходное значение меняется на
При данной неисправности может кратковременно помочь выключение зажигания с последующим повторным пуском двигателя, чтобы ЭБУ сохранил новое значение выходного сигнала, как положение закрытого дросселя.

Установить наличие данной неисправности датчика положения дроссельной заслонки можно путем измерения выходного значения на холодном двигателе (не работавшем не менее 2,5 часов) и на прогретом двигателе. Если значение сильно различаются имеет место быть данный дефект и датчик необходимо менять на более качественный.
Статья взята с сайта http://елм327.рф
Что такое ДПДЗ и как он ломается
- Главная
- Статьи
- Много бензина, мало тяги и нет холостых: что такое ДПДЗ и как он ломается
Автор: Михаил Баландин
Датчик положения дроссельной заслонки – не самый сложный датчик автомобиля. И ломается он, в общем-то, редко. Но если он выходит из строя, поиск неисправности может затянуться: датчик этот хоть и не сложный, но очень нужный.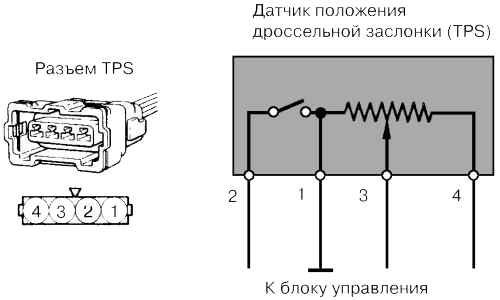
Есть контакт, нет контакта
Задача датчика положения дроссельной заслонки (или ДПДЗ) простая: передать в электронный блок управления двигателем (ЭБУ) данные о том, на какой угол (проще говоря, насколько сильно) открыта дроссельная заслонка. Каждый раз, когда водитель нажимает или отпускает педаль газа, заслонка меняет угол открытия, а значит, мотору нужно изменить количество подаваемого топлива (чтобы сохранить качество топливо-воздушной смеси, близкое к стехиометрической 14,7:1) и немного подкорректировать угол опережения зажигания. Проблемы со смесью и с углом опережения зажигания возникнут при любой проблеме с ДПДЗ: если сигнала от него не будет вовсе или если он будет неверным.
Так как датчику требуется подавать лишь одно значение, то электрическая часть у него примитивная: есть только одно выходное значение напряжения, которое изменяется в пределах приблизительно от 0,4 до 5 вольт в зависимости от угла поворота заслонки, и постоянное питающее напряжение от бортовой сети автомобиля для обеспечения работы датчика. Эта часть у всех ДПДЗ одинаковая.
Эта часть у всех ДПДЗ одинаковая.
А вот механическая часть может быть разной. Чаще выходит из строя контактный (или плёночно-резистивный) датчик. Причём основной источник проблемы – сам контакт. В датчике такого типа есть ползунок, который ходит по резистивным дорожкам (если проще, то по длинным дорожкам с большим сопротивлением). Работает эта штука по принципу переменного резистора: при перемещении ползунка меняется сопротивление, а значит, и выходное напряжение, которое является сигналом для ЭБУ. Простота – это, конечно, хорошо, но в случае с плёночно-резистивным датчиком именно эта простота часто и становится причиной его поломки. Точнее, износа механической части – самого датчика или дорожки. От постоянного движения они постепенно стачиваются, и в один не очень прекрасный момент контакт между ними или пропадает, или становится нестабильным. В первом случае сигнала на ЭБУ нет, во втором он есть, но неверный. Как правило, скачкообразный.
Плохо и первое, и второе, потому что для правильной работы ЭБУ требуется постоянно точное значение угла открытия заслонки. Например, открыта она на 12% – так и видит ЭБУ, а если водитель нажимает на газ и открывает заслонку, например, на 82%, то ЭБУ должен видеть весь процесс с плавным изменением напряжения сигнала ДПДЗ от соответствующего открытию в начале движения педали газа до соответствующего его окончанию. А в итоге он может увидеть, что сначала заслонка была открыта на те самые 12%, а потом резко – на 82. И в этом случае никакого разгона и тем более «подхвата» при нажатии на педаль газа не случится: нет изменения положения ДПДЗ – нет и топлива. Машина может дёргаться (если вдруг на каком-то участке резистивной дорожки вдруг появится контакт с ползунком), но нормально ехать не будет. В целом, описанное выше – это наиболее частая неприятность с ДПДЗ. Но, к сожалению, не единственная.
Например, открыта она на 12% – так и видит ЭБУ, а если водитель нажимает на газ и открывает заслонку, например, на 82%, то ЭБУ должен видеть весь процесс с плавным изменением напряжения сигнала ДПДЗ от соответствующего открытию в начале движения педали газа до соответствующего его окончанию. А в итоге он может увидеть, что сначала заслонка была открыта на те самые 12%, а потом резко – на 82. И в этом случае никакого разгона и тем более «подхвата» при нажатии на педаль газа не случится: нет изменения положения ДПДЗ – нет и топлива. Машина может дёргаться (если вдруг на каком-то участке резистивной дорожки вдруг появится контакт с ползунком), но нормально ехать не будет. В целом, описанное выше – это наиболее частая неприятность с ДПДЗ. Но, к сожалению, не единственная.
Реже в ДПДЗ такого типа может износиться сам привод ползунка. Почти экзотика, но бывает.
Второй тип датчиков положения заслонки – бесконтактный или магниторезистивный. Тут механического контакта нет, и внутри датчика ломаться нечему – там нет механики.
Вместе с тем у датчиков обоих типов есть общее слабое место – их проводка. Тут неисправности классические: либо обрыв, либо короткое замыкание. И разницы в способах проверки датчиков нет, но об этом чуть ниже. А пока поговорим о пользе сканера.
Ты не разберёшь, пока не повернёшь…
Чем плоха поломка ДПДЗ, так это списком сопутствующих проблем. Если износ ступичного подшипника, стойки стабилизатора и ещё кучи других деталей можно определить сразу, то с датчиком заслонки придётся повозиться. А что ещё делать, если есть десятки разных симптомов неисправности?
Наиболее очевидный признак неисправности ДПДЗ – это нестабильный холостой ход. Правда, он может быть настолько нестабильным, что мысль о датчике заслонки может прийти последней. Холостые могут просто плавать, а могут плавать настолько сильно, что мотор будет сам по себе глохнуть. Однако всё то же самое может быть и по огромному количеству других причин, начиная от «глюков» датчика положения коленвала до подсоса воздуха на впуске. Более того, остальные симптомы ясности тоже не добавят. А их множество: снижение мощности, рост расхода топлива, снижение динамики, провалы и рывки при разгоне и просто нестабильная (по ощущениям – «неправильная») работа двигателя. Никаких определённых выводов по этим признакам сделать невозможно, зато можно заменить половину машины, потратить много денег и остаться с той же неисправностью. Поэтому повторю ещё раз: лучше всего начинать ремонт с диагностики. Тем более что неисправности ДПДЗ очень легко найти сканером.
Более того, остальные симптомы ясности тоже не добавят. А их множество: снижение мощности, рост расхода топлива, снижение динамики, провалы и рывки при разгоне и просто нестабильная (по ощущениям – «неправильная») работа двигателя. Никаких определённых выводов по этим признакам сделать невозможно, зато можно заменить половину машины, потратить много денег и остаться с той же неисправностью. Поэтому повторю ещё раз: лучше всего начинать ремонт с диагностики. Тем более что неисправности ДПДЗ очень легко найти сканером.
Начать можно с поиска ошибок. В случае с ДПДЗ коды могут быть разными. Например, P0120, Р2135, P0122, P0123 , P0220, P0222 или P0223. Если какой-то из кодов в ошибках есть, желательно провести тест заслонки с контролем показаний датчика в онлайн-режиме. При изменении угла поворота заслонки должно быть заметно плавное изменение напряжение с сигнального контакта датчика и открытие заслонки в процентах. Если есть скачки, датчик придётся заменить.
К сожалению, сканер есть дома не у каждого, поэтому посмотрим, как другим способом можно проверить этот вредный ДПДЗ. Для этого другого способа понадобится простейший вольтметр (то есть любой мультиметр). Проверка пройдёт в три этапа.
Для этого другого способа понадобится простейший вольтметр (то есть любой мультиметр). Проверка пройдёт в три этапа.
На первом этапе нужно проверить, подаётся ли на ДПДЗ напряжение. Для этого снимаем разъём датчика и измеряем на нём напряжение. Удаётся найти подходящие 12-13 вольт – всё хорошо. Нет – надо искать, куда это напряжение делось. Возможно, в проводке датчика есть обрыв или короткое замыкание.
Если с напряжением всё в порядке, переходим ко второму этапу: смотрим крайние значения сигнала датчика. Для этого сначала ищем сигнальный контакт с датчика: подсоединяем минусовую клемму вольтметра к массе, плюсом ищем напряжение в 0,5-5 В на пине разъёма. Как только нужный пин найден (а это сделать несложно, там есть только разъёмы питания и сигнала), поворачиваем заслонку в крайние положения и следим за напряжением. При полностью закрытой заслонке напряжение должно быть около 0,4-0,6 В, при полностью открытой – больше 4 В. Смысл этой операции заключается в том, что очень часто в контактных датчиках резистивные дорожки изнашиваются в самом крайнем положении, соответствующем полностью закрытой заслонке – ползунок там находится в момент холостого хода, оттуда он каждый раз начинает своё движение при нажатии на педаль газа. Если в крайних положениях заслонки напряжение не выходит за границы нормы, переходим к третьему этапу.
Если в крайних положениях заслонки напряжение не выходит за границы нормы, переходим к третьему этапу.
На нём нам важно узнать, нет ли скачков данных с датчика. Хорошо, если есть сканер: там скачки можно увидеть даже не столько по цифрам, сколько по графику (если сканер имеет возможность увидеть эту информацию в режиме теста и вывести её в графическом виде). А вот с мультиметром придётся быть аккуратным и внимательным. Теоретически, конечно, ничего сложного нет: надо плавно поворачивать дроссельную заслонку и одновременно следить за напряжением, которое показывает вольтметр. Цифры должны меняться плавно, соответствуя повороту заслонки. Если есть скачки, то ползунок или дорожка имеют дефекты, а датчик работать нормально уже не может.
Что делать?
Датчик положения дроссельной заслонки – деталь надёжная и из строя выходит редко. Какого-то способа продлить ему жизнь не существует: механического износа контактного ДПДЗ избежать невозможно. Пытаться ремонтировать этот датчик – занятие неблагодарное. Проще купить новый, что обычно и делают.
Проще купить новый, что обычно и делают.
При этом надо помнить, что ездить с неисправным датчиком обычно хоть и можно, но сложно. Кроме того, неисправность ДПДЗ может привести к более серьёзным поломкам. Например, из-за неправильно выставленного блоком управления двигателя угла опережения зажигания или бедной смеси двигатель не только будет плохо тянуть, но может и перегреться. Появившиеся рывки при разгоне не понравятся трансмиссии, особенно автоматической коробке (любого типа). Ну а пойти на обгон и вдруг потерять тягу – это попросту опасно. Так что при появлении любых признаков неисправности ДПДЗ лучше не откладывать диагностику и ремонт в долгий ящик: отложенный на неопределённый срок ремонт обычно получается дороже своевременного.
практика
Новые статьи
Статьи / Практика
Вечная молодость: как не дать постареть кузову автомобиля
Те, кто хотя бы раз в жизни покупал новый автомобиль, испытал чувство, которое можно описать расхожей фразой «лишь бы с ласточкой ничего не случилось». Но жизнь так погано устроена, что с ла…
1712
0
4
27.12.2022
Но жизнь так погано устроена, что с ла…
1712
0
4
27.12.2022
Статьи / Популярные вопросы Зона действия дорожных знаков: как ее определить Дорожные знаки хорошо знают практически все водители. Но многие спустя годы после автошколы иногда задаются вопросом: «докуда действует вот тот знак, который я только что проехал?». В голову… 1234 0 1 27.12.2022
Статьи / Авто с пробегом
5 причин покупать и не покупать Kia Cerato III (YD)
Довольно большой, но недорогой седан в России не мог не понравиться покупателям. И Cerato понравился. Автомобили третьего поколения появились в 2013 году, и с каждым годом продажи только рос…
5353
3
1
25.12.2022
И Cerato понравился. Автомобили третьего поколения появились в 2013 году, и с каждым годом продажи только рос…
5353
3
1
25.12.2022
Популярные тест-драйвы
Тест-драйвы / Тест-драйв Haval Dargo против Mitsubishi Outlander: собака лает, чужестранец идет В дилерском центре Haval на юге Москвы жизнь кипит: покупатели разглядывают машины, общаются с менеджерами и подписывают какие-то бумаги. Пока я ждал выдачи тестового Dargo, такой же кроссов… 20422 7 205 13.09.2022
Тест-драйвы / Тест-драйв
Мотор от Mercedes, эмблема от Renault, сборка от Dacia: тест-драйв европейского Logan 1,0
Казалось бы, что нового можно рассказать про Renault Logan второго поколения, известный каждому российскому таксисту, что называется, вдоль и поперёк? Однако конкретно в этом автомобиле есть. ..
16202
10
41
13.08.2022
..
16202
10
41
13.08.2022
Тест-драйвы / Тест-драйв Geely Coolray против Haval Jolion: бесплатный сыр? Если бы! Хотите купить сегодня машину с полноценной гарантией, в кредит по адекватной ставке, без диких дилерских накруток? Сейчас это та еще задачка, ведь полноценную цепочку «представительство – з… 13678 26 30 10.08.2022
Опасности Delta P (перепада давления) при дайвинге
Опасности Delta P (перепада давления) при дайвинге Опасности при дайвинге, вызванные перепадом давления, возникают, когда вода перемещается из области высокого давления в область более низкого давления. Риска нет, когда нет потока воды, но как только поток начинается, прилагаемые силы обычно очень значительны.
Риска нет, когда нет потока воды, но как только поток начинается, прилагаемые силы обычно очень значительны.
Существует множество ситуаций, когда возникает поток воды, это может быть связано с открытием клапана, попаданием в пустоту или запуском насоса. Когда поток проходит через отверстие, любой дайвер, приближающийся к стороне высокого давления, может быть втянут и пойман в ловушку. В таких условиях часто возникают серьезные или смертельные травмы. Разница в высоте между двумя водоемами на разных уровнях создает всасывающую силу через любое отверстие или отверстие в преграде между ними. Чем больше разница в высоте или чем больше отверстие в барьере, тем больше и опаснее сила всасывания. Тем не менее, очень значительные силы всасывания могут также создаваться при небольшой разнице уровней воды и относительно большом открытии, это может быть смертельным даже на малых глубинах 3 метра и менее.
Перед началом водолазных работ необходимо провести соответствующие процедуры оценки рисков.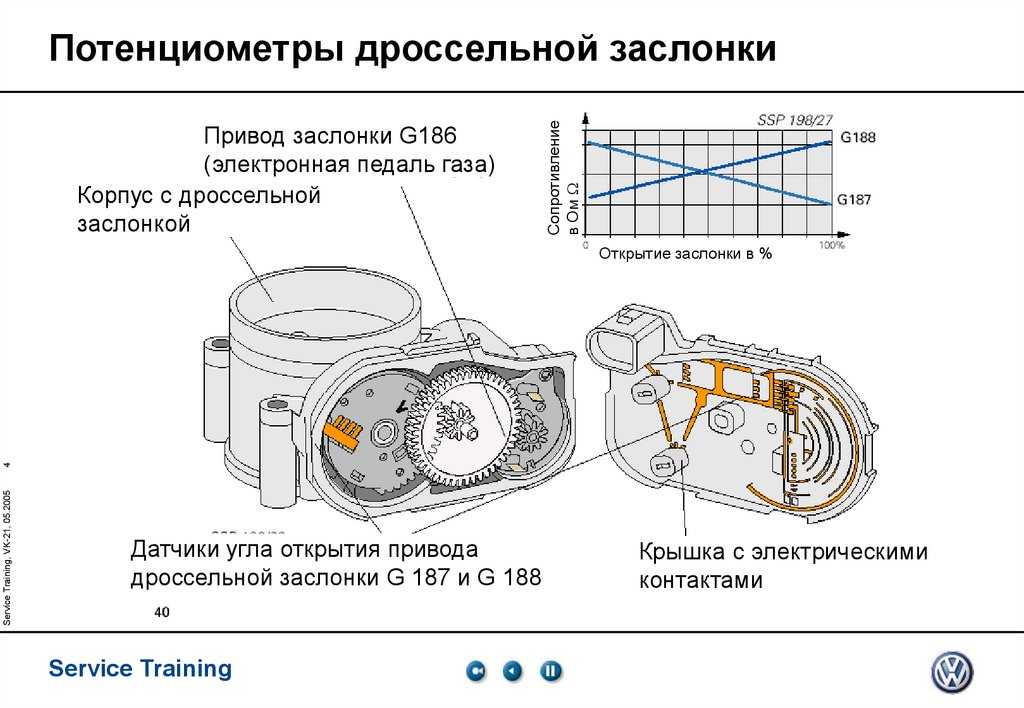 Перед любыми водолазными операциями, в которых выявлена потенциальная опасность Delta P, должны быть введены в действие процедуры изоляции и блокировки / маркировки. Всегда предпочтительнее иметь двойную изоляцию, где это возможно.
Перед любыми водолазными операциями, в которых выявлена потенциальная опасность Delta P, должны быть введены в действие процедуры изоляции и блокировки / маркировки. Всегда предпочтительнее иметь двойную изоляцию, где это возможно.
Особое внимание всегда следует уделять следующим местам:
• Насосы, заборники, клапаны и дренажи.
• Плотины, шлюзы, ворота и водосливы.
• Трещины/пустоты в трубах или шлангах.
• Гребные винты, подруливающие устройства на судах и вокруг них.
• Большое оборудование для подводного перемещения.
• Риск выхода из строя установки или оборудования.
• Затопленные трубы или пустые конструкции, загерметизированные при атмосферном давлении или давлении выше или ниже давления окружающей воды.
Расчет силы от перепада давления
Сила = Давление x Площадь
Сила (сила в килограммах) = Разница уровней x Плотность воды x Площадь
Плотность воды = 1025 кг/м 3 для морской воды
или 1000 кг/м 3 для пресной воды
Область = π x (диаметр/2) = 3,5 x 1025 x π x (0,3/2) = 254 кгс
Отношение скорости воды к расстоянию до опасного места Дайверы подвергаются опасности из-за потока воды, всасывания или турбулентности. Область быстро движущейся воды вокруг этих входов (будь то в результате работы или отказа оборудования и оборудования) называется зоной опасности перепада давления (DPDZ). Дайверы обычно не могут обнаружить вокруг себя опасные перепады давления, когда они погружены в воду, а как только они это сделают, будет уже слишком поздно.
Область быстро движущейся воды вокруг этих входов (будь то в результате работы или отказа оборудования и оборудования) называется зоной опасности перепада давления (DPDZ). Дайверы обычно не могут обнаружить вокруг себя опасные перепады давления, когда они погружены в воду, а как только они это сделают, будет уже слишком поздно.
Как показано на рисунке выше, дайвер, который движется от точки a к b , не почувствует изменения расхода воды, но при перемещении на то же расстояние, 300 мм, от b к c дайвер испытает экспоненциально сильное увеличение скорости потока и будет втянут.
В приведенных ниже таблицах приведены значения силы и расхода в трубах разного диаметра на разной глубине.
Сила перепада давления в трубе (кгс) Скорость потока воды в трубе (м/с) Морфогенез 1. Амано М., М. Ито, К. Кимура, Ю. Фуката, К. Чихара, Т. Накано, Ю. Мацуура и К. Кайбути. 1996. Фосфорилирование и активация миозина Rho-ассоциированной киназой (Rho-киназой). Дж. Биол. хим. 271 : 20246-20249. [PubMed] [Google Scholar]
Чихара, Т. Накано, Ю. Мацуура и К. Кайбути. 1996. Фосфорилирование и активация миозина Rho-ассоциированной киназой (Rho-киназой). Дж. Биол. хим. 271 : 20246-20249. [PubMed] [Google Scholar]
2. Аша Х., Н. Д. де Руйтер, М. Г. Ван и И. К. Харихаран. 1999. ГТФаза Rap1 функционирует как регулятор морфогенеза in vivo. EMBO J. 18 : 605-615. [Бесплатная статья PMC] [PubMed] [Google Scholar]
3. Baena-López, L.A., A. Baonza и A. Garcia-Bellido. 2005. Ориентация клеточных делений определяет форму органов дрозофилы. Курс. биол. 15 : 1640-1644. [PubMed] [Академия Google]
4. Боттнер Б., П. Харджес, С. Ишимару, М. Хеке, Х. К. Фан, Ю. Цинь, Л. Ван Элст и У. Галл. 2003. Гомолог AF-6 canoe действует как эффектор Rap1 во время дорсального закрытия эмбриона дрозофилы. Генетика 165 : 159-169. [Бесплатная статья PMC] [PubMed] [Google Scholar]
5. Бос, Дж. Л. 2005. Связь Rap с клеточной адгезией. Курс. мнение Клеточная биол. 17 : 123-128. [PubMed] [Google Scholar]
Бос, Дж. Л. 2005. Связь Rap с клеточной адгезией. Курс. мнение Клеточная биол. 17 : 123-128. [PubMed] [Google Scholar]
6. Бос, Дж. Л., К. де Брюйн, Дж. Энсеринк, Б. Куйперий, С. Рангараджан, Х. Реманн, Дж. Ридл, Дж. де Рой, Ф. ван Мансфельд и Ф. Зварткруйс. 2003. Роль Rap1 в интегрин-опосредованной клеточной адгезии. Биохим. соц. Транс. 31 : 83-86. [PubMed] [Google Scholar]
7. Bosgraaf, L., and PJ van Haastert. 2006. Регуляция миозина II у Dictyostelium. Евро. Дж. Клеточная биология. 85 : 969-979. [PubMed] [Google Scholar]
8. Брага В.М., Яп А.С. 2005. Проблемы изобилия: эпителиальные соединения и передача сигналов малых ГТФаз. Курс. мнение Клеточная биол. 17 : 466-474. [PubMed] [Google Scholar]
9. Брэнд, А. Х. и Н. Перримон. 1993. Направленная экспрессия генов как средство изменения клеточных судеб и создания доминантных фенотипов.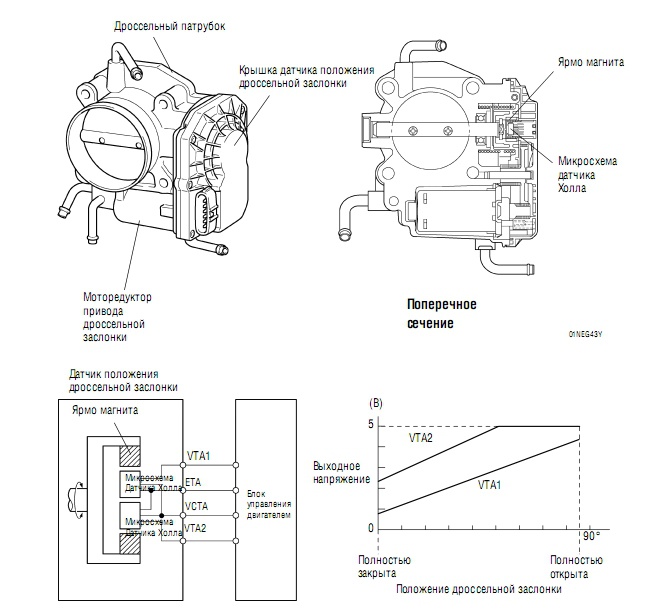 Развитие 118 : 401-415. [PubMed] [Google Scholar]
Развитие 118 : 401-415. [PubMed] [Google Scholar]
10. Чоу Т. Б. и Н. Перримон. 1996. Аутосомный метод FLP-DFS для создания мозаик зародышевой линии у Drosophila melanogaster. Генетика 144 : 1673-1679. [Бесплатная статья PMC] [PubMed] [Google Scholar]
11. Dawes-Hoang, R.E., K.M. Parmar, A.E. Christiansen, C.B. Phelps, A.H. Brand и E.F. Wieschaus. 2005. Складчатая гаструляция, изменение формы клеток и контроль локализации миозина. Развитие 132 : 4165-4178. [PubMed] [Google Scholar]
12. de Rooij, J., N. M. Boenink, M. van Triest, R. H. Cool, A. Wittinghofer, and J. L. Bos. 1999. PDZ-GEF1, фактор обмена гуаниновых нуклеотидов, специфичный для Rap1 и Rap2. Дж. Биол. хим. 274 : 38125-38130. [PubMed] [Google Scholar]
13. Франке Дж. Д., Р. А. Монтегю и Д. П. Кихарт. 2005. Немышечный миозин II генерирует силы, которые передают напряжение и вызывают сокращение во многих тканях во время дорсального закрытия. Курс. биол. 15 : 2208-2221. [PubMed] [Google Scholar]
Курс. биол. 15 : 2208-2221. [PubMed] [Google Scholar]
14. Гарсия-Беллидо, А., и Э. Б. Льюис. 1976. Автономная клеточная дифференцировка гомеотических bithorax мутантов Drosophila melanogaster. Дев. биол. 48 : 400-410. [PubMed] [Google Scholar]
15. Гарсия-Беллидо, А., П. Риполл и Г. Мората. 1976. Компартментализация дорсального мезоторакального диска дрозофилы. Дев. биол. 48 : 132-147. [PubMed] [Google Scholar]
16. Гибсон М.С. и Н. Перримон. 2003. Апикобазальная поляризация: эпителиальная форма и функция. Курс. мнение Клеточная биол. 15 : 747-752. [PubMed] [Google Scholar]
17. Хайго, С. Л., Дж. Д. Хильдебранд, Р. М. Харланд и Дж. Б. Уоллингфорд. 2003. Shroom вызывает апикальное сужение и необходим для формирования шарнирной точки во время закрытия нервной трубки. Курс. биол. 13 : 2125-2137. [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
18. Halsell, S. R., B. I. Chu, and D. P. Kiehart. 2000. Генетический анализ демонстрирует прямую связь между передачей сигналов rho и немышечной функцией миозина во время морфогенеза Drosophila. Генетика 155 : 1253-1265. [Бесплатная статья PMC] [PubMed] [Google Scholar]
19. Харихаран И. К., Р. В. Картью и Г. М. Рубин. 1991. Мутация Drosophila roughened: активация гомолога rap нарушает развитие глаз и препятствует детерминации клеток. Сотовый 67 : 717-722. [PubMed] [Google Scholar]
20. Hildebrand, JD 2005. Shroom регулирует форму эпителиальных клеток посредством апикального расположения актомиозиновой сети. Дж. Клеточные науки. 118 : 5191-5203. [PubMed] [Google Scholar]
21. Хильдебранд, Дж. Д., и П. Сориано. 1999. Shroom, актин-связывающий белок, содержащий домен PDZ, необходим для морфогенеза нервной трубки у мышей. Ячейка 99 : 485-497. [PubMed] [Google Scholar]
Ячейка 99 : 485-497. [PubMed] [Google Scholar]
22. Huelsmann, S., C. Hepper, D. Marchese, C. Knoll, and R. Reuter. 2006. Головокружение PDZ-GEF регулирует форму клеток мигрирующих макрофагов посредством Rap1 и интегринов у эмбрионов дрозофилы. Развитие 133 : 2915-2924. [PubMed] [Google Scholar]
23. Jamora, C., and E. Fuchs. 2002. Межклеточная адгезия, передача сигналов и цитоскелет. Нац. Клеточная биол. 4 : Е101-Е108. [PubMed] [Академия Google]
24. Чон, Т.Дж., Д.Дж. Ли, С. Мерло, Г. Уикс и Р.А. Фиртел. 2007. Rap1 контролирует клеточную адгезию и подвижность клеток посредством регуляции миозина II. Дж. Клеточная биология. 176 : 1021-1033. [Бесплатная статья PMC] [PubMed] [Google Scholar]
25. Кальчмидт, Дж. А., Н. Лоуренс, В. Морель, Т. Балайо, Б. Г. Фернандес, А. Пелисье, А. Хасинто и А. Мартинес Ариас. 2002. Планарная полярность и динамика актина в эпидермисе дрозофилы. Нац. Клеточная биол. 4 : 937-944. [PubMed] [Google Scholar]
Планарная полярность и динамика актина в эпидермисе дрозофилы. Нац. Клеточная биол. 4 : 937-944. [PubMed] [Google Scholar]
26. Кавадзири А., Н. Ито, М. Фуката, М. Накагава, М. Ямага, А. Ивамацу и К. Кайбути. 2000. Идентификация нового белка, взаимодействующего с бета-катенином. Биохим. Биофиз. Рез. коммун. 273 : 712-717. [PubMed] [Google Scholar]
27. Келлер Р., Л. А. Дэвидсон и Д. Р. Шук. 2003. Как мы устроены: биомеханика гаструляции. Дифференциация 71 : 171-205. [PubMed] [Академия Google]
28. Kiehart, DP 1990. Молекулярно-генетическое исследование функции тяжелой цепи миозина. Сотовый 60 : 347-350. [PubMed] [Google Scholar]
29. Кихарт Д. П., К. Г. Гэлбрейт, К. А. Эдвардс, У. Л. Риколл и Р. А. Монтегю. 2000. Множественные силы способствуют морфогенезу клеточного листа для закрытия спины у Drosophila.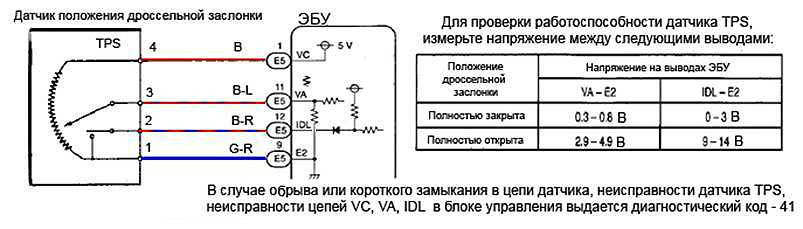 Дж. Клеточная биология. 149 : 471-490. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Дж. Клеточная биология. 149 : 471-490. [Бесплатная статья PMC] [PubMed] [Google Scholar]
30. Кимура К., М. Ито, М. Амано, К. Чихара, Ю. Фуката, М. Накафуку, Б. Ямамори, Дж. Фэн, Т. Накано, К. Окава, А. Ивамацу и К. Кайбути. 1996. Регуляция миозинфосфатазы с помощью Rho и Rho-ассоциированной киназы (Rho-киназы). Наука 273 : 245-248. [PubMed] [Google Scholar]
31. Нокс А. Л. и Н. Х. Браун. 2002. Регуляция Rap1 GTPase позиционирования соединения адгезивов и клеточной адгезии. Наука 295 : 1285-1288. [PubMed] [Google Scholar]
32. Kooistra, M. R., N. Dube, and J. L. Bos. 2007. Rap1: ключевой регулятор образования межклеточных соединений. Дж. Клеточные науки. 120 : 17-22. [PubMed] [Google Scholar]
33. Кеппен М., Б. Г. Фернандес, Л. Карвалью, А. Хасинто и С. П. Гейзенберг. 2006. Скоординированные изменения формы клеток контролируют движение эпителия у рыбок данио и дрозофилы.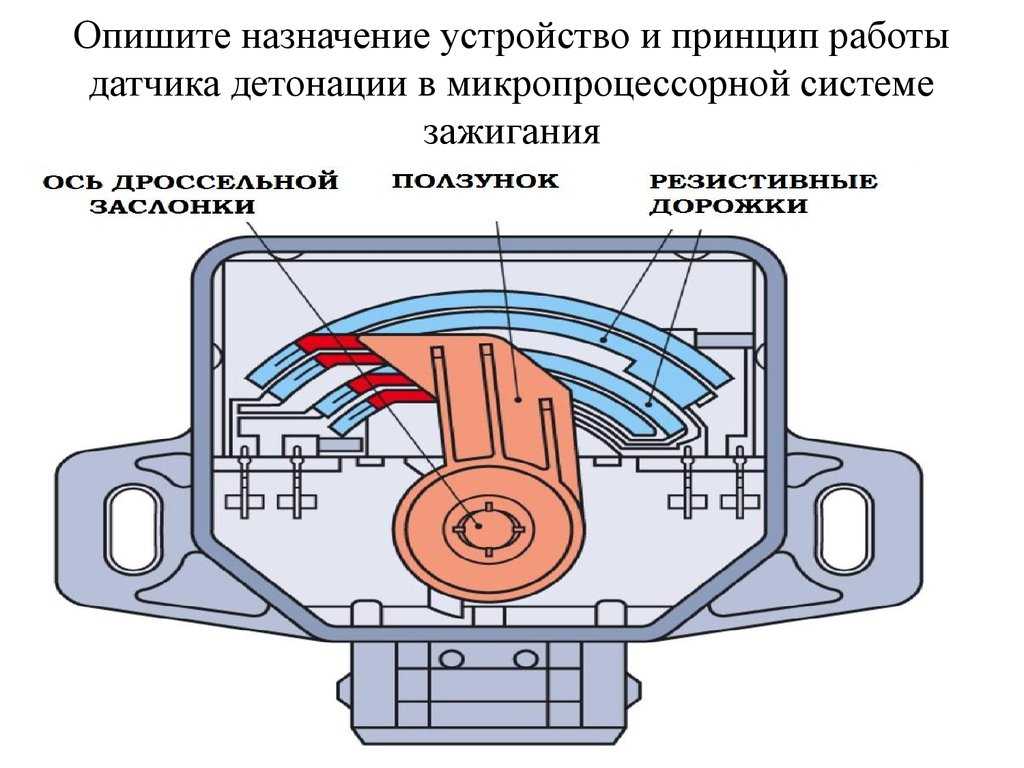 Развитие 133 : 2671-2681. [PubMed] [Google Scholar]
Развитие 133 : 2671-2681. [PubMed] [Google Scholar]
34. Кругманн С., Р. Уильямс, Л. Стивенс и П. Т. Хокинс. 2004. ARAP3 представляет собой GAP, регулируемый PI3K и rap для RhoA. Курс. биол. 14 : 1380-1384. [PubMed] [Академия Google]
35. Ли, Дж. Х., К. С. Чо, Дж. Ли, Д. Ким, С. Б. Ли, Дж. Ю, Г. Х. Ча и Дж. Чанг. 2002. Drosophila PDZ-GEF, фактор обмена гуаниновых нуклеотидов на Rap1 GTPase, обнаруживает новый вышестоящий регуляторный механизм в сигнальном пути митоген-активируемой протеинкиназы. Мол. Клетка. биол. 22 : 7658-7666. [Бесплатная статья PMC] [PubMed] [Google Scholar]
36. Lee, YS, and RW Carthew. 2003. Создание лучшего вектора РНК-интерференции для дрозофилы: использование интронных спейсеров. Методы 30 : 322-329. [PubMed] [Google Scholar]
37. Luo, L., YJ Liao, LY Jan, and YN Jan. 1994. Различные морфогенетические функции сходных малых GTPases: Drosophila Drac1 участвует в разрастании аксонов и слиянии миобластов. Гены Дев. 8 : 1787-1802. [PubMed] [Google Scholar]
Гены Дев. 8 : 1787-1802. [PubMed] [Google Scholar]
38. Major, RJ, and KD Irvine. 2006. Локализация и потребность в миозине II на границе дорсально-вентрального отдела крыла дрозофилы. Дев. Дин. 235 : 3051-3058. [PubMed] [Google Scholar]
39. Мартин П. и С. М. Паркхерст. 2004. Параллели между восстановлением тканей и морфогенезом эмбриона. Развитие 131 : 3021-3034. [PubMed] [Google Scholar]
40. Мино А., Т. Оцука, Э. Иноуэ и Ю. Такай. 2000. Мембран-ассоциированная гуанилаткиназа с инвертированной ориентацией (MAGI)-1/мозговой ингибитор ангиогенеза 1-ассоциированный белок (BAP1) в качестве каркасной молекулы для Rap small G-белка GDP/GTP обменного белка в плотных соединениях. Гены Клетки 5 : 1009-1016. [PubMed] [Google Scholar]
41. Мишра С., Смолик С.М., Форте М.А., Сторк П.Дж. 2005. Независимая от Ras активация передачи сигналов ERK через тирозинкиназу рецептора туловища опосредуется Rap1. Курс. биол. 15 : 366-370. [PubMed] [Google Scholar]
Курс. биол. 15 : 366-370. [PubMed] [Google Scholar]
42. Мората Г. и П. Риполл. 1975. Минуты: мутанты дрозофилы, автономно влияющие на скорость деления клеток. Дев. биол. 42 : 211-221. [PubMed] [Академия Google]
43. Николаиду К.К. и К. Барретт. 2004. Сигнальный путь Rho GTPase повторно используется в укладке эпителия и потенциально выбирает результат активации Rho. Курс. биол. 14 : 1822-1826. [PubMed] [Google Scholar]
44. Палади М. и У. Тепасс. 2004. Функция Rho GTPases в эмбриональной миграции клеток крови у дрозофилы. Дж. Клеточные науки. 117 : 6313-6326. [PubMed] [Google Scholar]
45. Патель, П. Х., Н. Тапар, Л. Гуо, М. Мартинес, Дж. Марис, К. Л. Гау, Дж. А. Ленгьел и Ф. Таманой. 2003. Drosophila Rheb GTPase необходима для прогрессирования клеточного цикла и роста клеток. Дж. Клеточные науки. 116 : 3601-3610.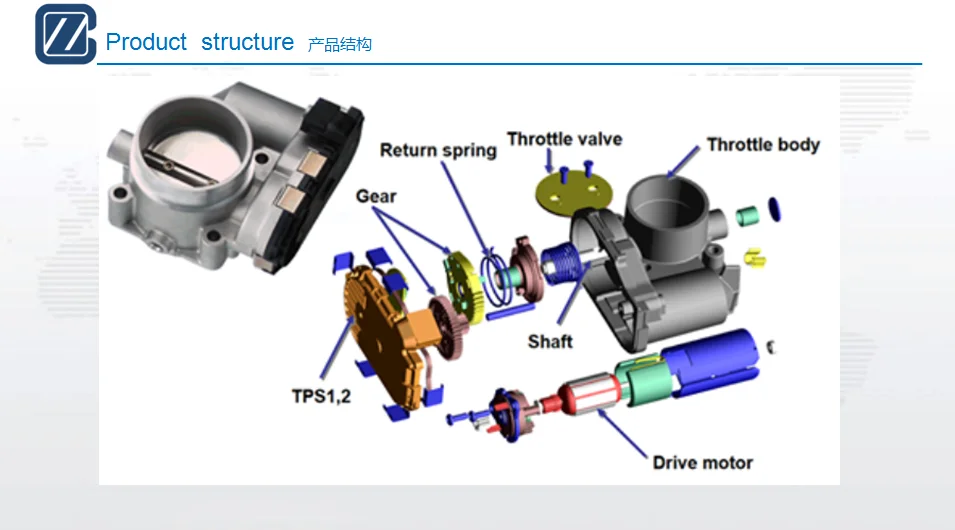 [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
46. Пеллис-ван Беркель В., М. Х. Верхейен, Э. Куппен, М. Асахина, Дж. де Рой, Г. Янсен, Р. Х. Пластерк, Дж. Л. Бос и Ф. Дж. Зварткруйс. 2005. Требование Caenorhabditis elegans RapGEF pxf-1 и rap-1 к целостности эпителия. Мол. биол. Сотовый 16 : 106-116. [Бесплатная статья PMC] [PubMed] [Google Scholar]
47. Савада Ю., Тамада М., Дубин-Талер Б.Дж., Чернявская О., Сакаи Р., Танака С. и Шитц М.П. 2006. Определение силы путем механического удлинения субстрата киназ семейства Src p130Cas. Сотовый 127 : 1015-1026. [Бесплатная статья PMC] [PubMed] [Google Scholar]
48. Spradling, AC, and GM Rubin. 1982. Транспозиция клонированных Р-элементов в хромосомы зародышевой линии дрозофилы. Science 218 : 341-347. [PubMed] [Академия Google]
49. Steimle, P.A., S. Yumura, G.P. Cote, Q.G. Medley, M.V. Polyakov, B. Leppert, and T. T. Egelhoff. 2001. Привлечение киназы тяжелой цепи миозина к богатым актином выпячиваниям Dictyostelium. Курс. биол. 11 : 708-713. [PubMed] [Google Scholar]
T. Egelhoff. 2001. Привлечение киназы тяжелой цепи миозина к богатым актином выпячиваниям Dictyostelium. Курс. биол. 11 : 708-713. [PubMed] [Google Scholar]
50. Такахаши К., Т. Мацуо, Т. Кацубэ, Р. Уэда и Д. Ямамото. 1998. Прямое связывание между двумя доменными белками PDZ Canoe и ZO-1 и их роль в регуляции N-концевого киназного пути jun в морфогенезе дрозофилы. мех. Дев. 78 : 97-111. [PubMed] [Google Scholar]
51. Тамада М., Т. Д. Перес, У. Дж. Нельсон и М. П. Шитц. 2007. Два различных режима сборки и динамики миозина во время закрытия эпителиальной раны. Дж. Клеточная биология. 176 : 27-33. [Бесплатная статья PMC] [PubMed] [Google Scholar]
52. Тамада М., М. П. Шитц и Ю. Савада. 2004. Активация сигнального каскада растяжением цитоскелета. Дев. Сотовый 7 : 709-718. [PubMed] [Академия Google]
53. Tepass, U. 1996. Крошки, компонент апикальной мембраны, необходимы для образования прилипателей zonula в первичном эпителии дрозофилы. Дев. биол. 177 : 217-225. [PubMed] [Google Scholar]
1996. Крошки, компонент апикальной мембраны, необходимы для образования прилипателей zonula в первичном эпителии дрозофилы. Дев. биол. 177 : 217-225. [PubMed] [Google Scholar]
54. Van Aelst, L. 1998. Двухгибридный анализ взаимодействий Ras-Raf. Методы Мол. биол. 84 : 201-222. [PubMed] [Google Scholar]
55. Verdier, V., GC Chen, and J. Settleman. 2006. Rho-киназа регулирует морфогенез тканей через немышечный миозин и LIM-киназу во время развития дрозофилы. BMC Дев. биол. 6 : 38. [Бесплатная статья PMC] [PubMed] [Google Scholar]
56. Уоллингфорд, Дж. Б., С. Э. Фрейзер и Р. М. Харланд. 2002. Конвергентное удлинение: молекулярный контроль движения поляризованных клеток во время эмбрионального развития. Дев. Сотовый 2 : 695-706. [PubMed] [Google Scholar]
57. Wang, H., S.R. Singh, Z. Zheng, S.W. Oh, X. Chen, K. Edwards, and S. X. Hou. 2006. Передача сигналов Rap-GEF контролирует прикрепление стволовых клеток к своей нише посредством регуляции DE-кадгерин-опосредованной клеточной адгезии в семенниках дрозофилы. Дев. Ячейка 10 : 117-126. [PubMed] [Google Scholar]
Edwards, and S. X. Hou. 2006. Передача сигналов Rap-GEF контролирует прикрепление стволовых клеток к своей нише посредством регуляции DE-кадгерин-опосредованной клеточной адгезии в семенниках дрозофилы. Дев. Ячейка 10 : 117-126. [PubMed] [Google Scholar]
58. Wei, S.Y., LM Escudero, F. Yu, LH Chang, LY Chen, YH Ho, CM Lin, C.S. Chou, W. Chia, J. Modolell, and JC Hsu. 2005. Эхиноид является компонентом слипчивых соединений, который взаимодействует с DE-кадгерином, опосредуя клеточную адгезию. Дев. Клетка. 8 : 493-504. [PubMed] [Google Scholar]
59. Winter, C.G., B. Wang, A. Ballew, A. Royou, R. Karess, JD Axelrod и L. Luo. 2001. Drosophila Rho-associated kinase (Drok) связывает Frizzled-опосредованную передачу сигналов планарной клеточной полярности с актиновым цитоскелетом. Сотовый 105 : 81-91. [PubMed] [Google Scholar]
60. Сюй Т. и Г. М. Рубин. 1993.